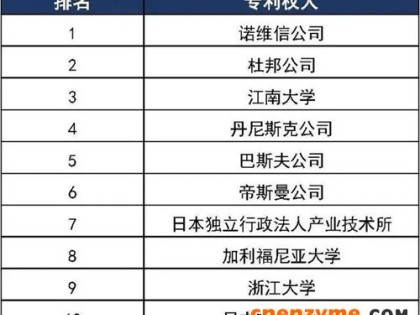
面包酵母羰基还原酶基因同源表达产物对映体ee 值和产率均有提高。近平滑假丝酵母基因组中克隆出新型 (S)-型羰基还原酶,重组菌 E. coliBL 21 能制备 (S)-苯基乙二醇,光学纯达 99.1%,产率为 89.6%[8,11]。
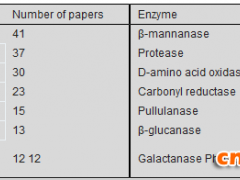
β-葡萄糖苷酶是纤维素酶系的重要组分,可将纤维二糖、可溶性纤维寡糖水解成葡萄糖及相应配基,水解 β-糖苷键及合成新的糖衍生物
。瑞士木霉 β-葡萄糖苷酶,通过基因敲除、氨基酸突变,突变体酶活力比野生型提高了 143 倍[9]。新鞘氨醇杆菌 β-葡萄糖苷酶基因在 E. coli 成功表达,可转化异黄酮糖苷生成相对应的苷元[10]。台湾乳白蚁肠道中的 β-葡萄糖苷酶,用原核表达系统进行异源表达,突变酶较野生型酶活力提高了 2.6 倍。亦提高了葡萄糖耐受性及热稳定性[10]。斜卧青霉 β-葡萄糖苷酶进行基因改造,通过转化获得 3 个表达菌株,酶活力比原始菌株提高 3.4–3.7 倍[10],从非解朊栖热菌分离到耐热 β-葡萄糖苷酶基因在 E. coli 中克隆表达,酶活力提高 17 倍[4]。从海洋宏基因组筛选到葡萄糖耐受的β-葡萄糖苷酶克隆到E. coli高效表达,葡萄糖浓度低于 400 mmol/L,对酶有促进作用,达到 1 000 mmol/L 葡萄糖浓度,酶活性为50%[8,11]。利用 DNA 改组和定点突变提高 β-葡萄糖苷酶稳定性,4 个突变体与野生型相比,61 ℃热失活半衰期提高了 14.16、68 和 44 倍。耐热性有明显提高[8]。
半乳糖苷酶,又名乳糖酶,能将乳糖分解成葡萄糖和半乳糖基,可以合成低聚糖。通过宏基因组法从海底泥中获得高效转糖基的 β-半乳糖苷酶,在 E. coli 中可溶性表达,对有机溶剂有耐受性,合成低聚半乳糖产量可达 51.6%[9]。曲霉 α-半乳糖苷酶固体发酵,酶活力达 305 IU/g[6]。硫磺矿硫化叶菌 Sulfolobus solfataricus P2 的 β-半乳糖苷酶进行分子改造构建了突变体,突变酶 F441Y 生产低聚半乳糖,产率达 61%,较野生型高 10%左右[9]。
腈水合酶在酰胺、羧酸及其衍生物合成中有重要应用价值,催化丙烯腈生产丙烯酰胺,诺卡氏菌 Nocardia sp. YS-2002 发酵活力最高达6 000 国际单位,在 E. coli 及毕赤酵母中得到表达[5]。
菊粉酶是水解菊粉的 β-2,1-D-果聚糖果糖苷键的水解酶,产低聚果糖,分内切酶和外切酶两种。将黑曲霉菊粉内切酶基因克隆到毕赤酵母得到高效表达,重组酶活力达 128 U/mL,达到国际水平[9]。
海藻糖合成酶,海藻糖在食品、医学、轻工业领域广泛应用,可用单酶法从麦芽糖生产海藻糖,通过酶分子的理性设计和 DNAshuffling 技术从两个 GRAS 菌种和两个嗜热菌中克隆到海藻糖合成酶基因并成功进行了高效表达,已实现了工业化生产[5]。
中性蛋白酶广泛用于工业化生产,枯草芽胞杆菌 AS1398 基因 Npr 的重组质粒转入 B. subtilisAS 1398,获得多拷贝的重组菌,连续传代
30代,重组菌质粒稳定性保持 100%,蛋白酶表达水平提高 1 倍以上,达到 24 000–28 000 U/mL[10]。有高效杀线虫毒力的侧孢短芽胞杆菌株经优化培养获得蛋白酶的活力达 12 379 U/mL,比优化前提高 5 倍,应用 B. subtilis WB600 表达体系,构建随机突变文库,获得热稳定的突变体,为生物杀虫剂提供依据[8]。人基质金属蛋白酶与肿瘤转移关系密切,在 E. coli 中成功克隆表达了该酶,为研究该酶与肿瘤转移机理和寻找该酶特异抑制剂奠定基础[5]。
葡聚糖酶属水解酶,分内切酶和外切酶,用于啤酒生产和饲料添加。嗜热青霉 β-1,3-1,4-葡聚糖酶基因克隆到原核表达载体并转入E.
coli BL 21 诱导表达,酶活力 240 U/mL[8],链霉菌S27内切β-1,3-葡聚糖酶基因在E. coli中高效表达,可水解昆布多糖等,可抑制致病真菌和产毒素真菌[8]。长梗木霉内切葡聚糖酶基因导入毕赤酵母表达,重组菌Ⅲ酶活力达 110 U/mL,优化后提高 50%[8]
。点突变极耐热 Thermotogamaritima 内切葡聚糖酶 Cell-12B,酶最适温度95 ℃,90 ℃仍保留 70%活力
[10]
。
N-乙酰神经氨酸是生物体内一种最重要的唾液酸,唾液酸糖链参与许多生命过程,CMP-唾液酸能促进神经细胞再生,对 CMP-唾液酸合成酶基因进行碱基改造,在 E. coli 中得到高效表达,酶表达量占总蛋白 26.5%,酶活力为 100 U/L,为出发菌株的 850 倍[4]。
黄曲霉毒素解毒酶为一种加氧酶,用重组技术构建基因在毕赤酵母中高密度发酵表达,酶占总蛋白量 56%,表达量达 814.5 mg/L[6]。将酶突变基因重组质粒引入 E. coli 扩增转入酿酒酵母,建立突变文库,突变体 A1773 酶活提高5 倍。突变体 A1242 耐高温性提高了 3.5 倍。突变体 DS1474 酶比活力 56 U/mg,突变体 DS896酶比活力为 44 U/mg[9],已获准在饲料中作为除毒的酶产品。
烟曲霉真菌感染是困扰人类健康的难题,系统研究烟曲霉基因组中 50 多个糖基化途径相关基因,先后从烟曲霉中克隆了几丁质酶、磷酸甘露糖异构酶、O-甘露糖转移酶、α-1,4-N-乙酰氨基葡萄糖转移酶、α-葡萄糖苷酶 I 及 CMP-唾液酸合成酶等基因、对了解真菌糖基化机制、开发基因工程药物、抗真菌感染药物有意义[6]。
L-天冬酰胺酶有明确治疗白血病的效果,通过 DNA 重组技术,将 L-天冬酰胺酶特异单链抗体片段与酶构建融合蛋白,提高了酶在体内的稳定性,重组 E. coli AS 1357 工程酶活力提高达 228 U/mL,相当野生菌的 50 多倍[4]。
用宏基因组学法将酯酶基因转入E. coli中,构建的工程菌提高了活力,表达量为 200 mg/L,酶活力达 180 U/mg,与野生型比较 60 ℃热稳定性提高 144 倍和 196 倍,获得了可降解氯氰菊酯、溴氰菊酯、氰戊菊酯和氟氯氰菊酯的新型聚酯类农药降解突变酯酶[9]。
重组毕赤酵母发酵生产碱性果胶酶,1 t 发酵罐产酶最高达 1 315U/mL[8]。黑曲霉 EIM-6 培养优化后果胶酶活力可提高至 30 231U/mL[8,11]。
假单胞菌水杨酸羟化酶基因克隆到 E. coli表达了两个萘降解酶,可降解水杨酸、乙酰水杨酸、磺基水杨酸、水杨醛、间硝基苯甲醛、5-氯水杨酸、辛醛及邻硝基苯甲醛等[5]。
有机磷酯类是一类高毒化合物,用于杀虫剂、除草剂或作为神经毒剂。克隆了假单胞菌有机磷酯水解酶,通过氨基酸位点突变,在 E. coli 中重组表达,对有机磷酯水解能力提高了约 7 000 倍[10]。