1. 微藻生物学研究
微藻是光合自养微生物,可以把CO2 和水转化为脂肪、碳水化合物等大分子有机物。在恶劣生长环境中(如氮饥饿),微藻体内能量主要以三酰甘油(TAGs)的形式贮藏。某些种类的微藻具有高效的光合作用和TAGs 积累能力(三酰甘油含量可占到干重的30-60%),油脂生产潜力巨大远远超过了传统的陆生植物。藻类的基础生物学研究是藻类筛选、培养及燃料转化等各项工艺的基础。微藻的基础生物学研究针对如何提高微藻油脂产率展开,具体方向有藻类光合作用、代谢途径及对模式生物的基因组学和系统生物学的研究。
1.1 光合作用
目前估计的微藻理论最高产量大致为100-200g-1m-2day-1,但微藻的确切理论最大产量是多少却没有一致的看法,造成伪造理论产量估算结果差距较大的部分原因是由于微藻培养物的透光、反射和吸收等参数的影响;另一个问题是在计算光合反应器产率时,通常只考虑反应器本身,而不考虑反应器所处的地理位置。理论产量是一个很重要的参数,可以以此设定微藻培养工艺的目标和菌株工程改造提高的目标。对微藻的光合作用机制的研究有助于提高微藻生物质产量,也会提高微藻的油脂含量。研究显示,部分藻类在限制光照时油脂含量会提高。本领域的研究重点将是光合作用与微藻脂质合成与含量的关系。
1.2 代谢途径
1.2.1 研究现状
微藻油脂产量不能达到理论值也说明我们目前对微藻中三酰甘油的合成机制、碳代谢途径和碳流分配缺乏清晰的认识。
随着代谢流分析方法的飞速发展,代谢物组学在系统水平得到进一步发展,在微生物系统方面,可以实现通过基因组、转录组数据、代谢途径分析和预测模型重建微生物代谢网络,藻类生物体中许多代谢途径及参与代谢的酶类已经得到了详细阐述,但对脂质中的碳流和碳分配还知之甚少。
在植物和微藻中,淀粉都是普通的碳和能量贮藏物质,与脂质三酰甘油有共同的合成前体,因此有互相转化的可能。已有研究表明,某些植物在脂质累积期之前,淀粉的合成和代谢是最为活跃的代谢途径,即淀粉是很重要能量储存物质。
最近有研究表明,如果高等植物中的淀粉合成途径被阻断或者受到抑制,植物胚芽或种子中的油脂累积量会降低40%。这些都表明淀粉(碳水化合物)合成与油脂合成之间有紧密的联系。但其相互作用实质/性质还是未知的。
1.2.1 重要研究方向
光合作用中,微藻细胞如何控制碳的流动和碳在不同细胞器和主要大分子之间(如碳氢化合物、蛋白质和脂质)的分配。
控制碳在脂质和其他储存性产物中分配的调控网络。
对脂质代谢和淀粉代谢间关系的研究,研究内容为微藻细胞内淀粉的合成、降解及如何参与脂质代谢。
1.3 基因组学和系统生物学
目前对微藻中脂质、碳水化合物等物质的合成和调节机制的了解非常有限,而要实现微藻的大规模培养,必须对微藻的生长特性有深入了解,需要借助基因组,代谢物组学、脂质组学等高通量分析工具,同时详细分析研究细胞的不同代谢途径。
1.3.1 模式生物体的选择
由于微藻种族发育的多样性,有大量模式系统可供研究。但实际上只能对有限的几种进行深入研究。考虑因素除了藻类合成的燃料前体外,还有分子和生化技术操作的难易程度、转录能力等。
1.3.1.1 蓝细菌
蓝细菌细胞中虽然不含有油脂,但是可以产生大量碳水化合物和次级代谢产物,生长速度快,既可以固氮又可以产氢,是颇具潜力的生物燃料原料。蓝细菌与绿藻相比,最大的优点是基因操作简单,可进行系统的基因学分析并对代谢途径进行改造。
为使蓝细菌中碳源的分配对燃料生产有利,必须移除一些储存碳的化合物,并从细胞和分子水平研究这种操作对生物体的影响。在不同的压力环境下会有不同蓝细菌中会累积不同的碳化合物,通过对不同突变体的系统生物学研究,提高研究人员对碳代谢流的理解,进一步设计生物燃料生产生物体。
模式蓝细菌Synechocystis sp. PCC6803 有望成为氢气生产中碳代谢流研究的模式生物体。除了Synechocystis sp. PCC6803 外,其它几株具有出色遗传系统的菌株也在研究中, 如Synechococcus7002 的产氢和产乙醇的研究,Anabaena7120 的产氢研究等。
1.3.1.2 绿藻
研究广泛的绿藻模式藻株有Chlamydomonas reinhardtii,Chlorella,Dunaliella salina,等。
Chlamydomonas reinhardtii 是研究最为深入的原核藻类,已经完成基因组测序,转基因技术也相当成熟。叶绿体转导技术已经建立,叶绿体蛋白的表达系统也很稳定,没有实现核转导技术,且大部分基因都在细胞核中,这可能成为其分析发展的一个障碍。
Chlorella 是另一种研究深入的绿藻,其中许多藻株都富含油脂。C.protothecoides 在培养基中添加碳源时,可以诱导异源生长,生长速率和油脂产量都会提高,最终油脂产量可以达到干重的50%以上。
Dunaliella salina 有一些利于大规模生物燃料生产的特征。它能够产生丰富的油脂,耐盐(可生长在0.1M 到饱和盐溶液中),在这种极端环境中也减少了其它生物体污染的几率,目前研究针对Dunaliella salina 进行基因测序工作,也有相关基因改造的报道。
1.3.1.3 硅藻
硅藻是美国水生计划中研究的主要藻种,油脂含量高,适用于生物燃料生产。硅藻环境适应性强,并且固定了全球20%的碳。
硅藻的特点是其细胞壁中含有二氧化硅,培养基中需要含有硅作为其营养元素。硅元素的缺乏是硅藻脂质累积的诱导因素之一,因为微藻的硅代谢途径与其他营养元素的代谢没有偶联,也没有参与到细胞内大分子的合成中,因此以硅饥饿诱导脂质累积比其它方法要简单。
目前,已经有两株硅藻完成了基因组测序,另外四株的测序工作正在进行中,但这几项测序计划的研究焦点不是生物燃料。部分硅藻的转基因技术已经成熟,基因表达的调控元素也已经鉴定出来。随着基因沉默技术和同源同组技术的发展,很快就会研制出硅藻操作工具盒。
1.3.2 模式生物体选择标准
1.3.2.1 系统生物学发展的模式系统选择标准
目前两种模式系统供选择:1.可反映燃料合成前体的细胞内调节反应基础信息的藻种和藻株。2.具有规模化培养潜力的藻种和藻株。
选择具体藻种时需要考虑以下因素:
藻种产生的燃料类型/副产物,可能的燃料产物包括H2、油脂、类异戊二烯、
碳水化合物、乙醇、甲醇等。
产物/产物中间体的分泌方式,如果藻种产生的燃料(前体)为细胞外分泌,可以省去藻种收获这一步骤。当然这也有一些问题,比如必须使用密闭式光合反应器。关于大规模生产中不同分泌类型的优缺点还需要进一步论证。
过程工艺相关特性。
异养/混合营养生长方式,在培养基中加入碳源后,部分藻种的油脂累积会有所提高,但细胞对碳源的利用方式还不清楚。如果微藻可以利用碳源,就可以
实现在光照条件下,和黑暗中都能够生长。缺点是:培养基中添加碳源会增加培养基被其他微生物污染的几率。
1.3.2.2 基因测序藻类筛选条件
基因组大小和重复片段。
种属研究的必要性。
根据路线图建议基因组研究的藻种有:Chlorella sp., Dunaliella sp.,
Nannochloropsis sp., Scenedesmus sp., Chlorococcum sp.,Peudochlorococcum sp.和部分硅藻。另外,采用生物信息学手段对获得的基因组序列进行分析基因组研究是,需要设立标准,确保生物信息学分析的一致性和可靠性。
2. 微藻筛选技术
2.1 优良藻种的保存
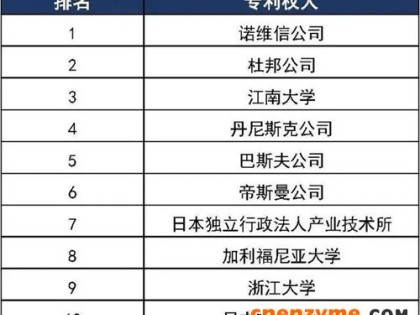
生产生物质燃料,优良藻种的获取至关重要。筛选出可用于规模化生产的高产、高品质的藻种,重点在于从自然界中直接分离筛选到新的原始藻株。世界上多个实验室已经筛选到大量藻种,并建立了藻种库,如UTEX 保藏有约3000 种藻种,CCMP 保藏藻种大于2500 种。但由于这些藻种已经培养了几十年,可能会发生变异,丢失一些原始性状。
2.2 藻种筛选条件
1)微藻生长在各种自然水域中,大部分商业化微藻生产设备都是开放式的
水池,生物质的获取依赖于藻类的自然繁殖。因此,在取样和筛选时,要考虑到微藻在自然条件中的繁殖能力。
2)大规模的取样与育种研究中,要筛选适应不同环境的藻种,以实用生产中对不同培养环境、代谢途径和代谢产物的需要。
3)除了考虑微藻的空间分布外,还要考虑其时间、季节性分布变化,另外,在同一水域中,既有浮游微藻,又有附着在水底的微藻,可应用在不同的培养过程中。因此路线图中也建议筛选样品要包括浮游藻类和附着在海底的藻类。
总之,理想的筛选过程需要考虑到生长生理学、产生的代谢物和藻株稳定性三个方面。
2.3 存在问题
1)筛选方法
传统的微藻筛选使用富集培养法,在分离单一菌株时要花费几周甚至数月的时间,筛选到的一些单细胞菌落有时是已经在培养的菌株。因此在大规模筛选菌株时,需要发展高通量自动筛选技术,荧光活化细胞分拣法(FACS)是一种高效筛选方法。
微藻大规模筛选的瓶颈在于缺乏可以同时兼顾多种因素的高通量筛选技术,和缺乏对藻株稳定性的测试环节。
藻株稳定性包括多种因素,如培养物的可持续性、环境适应性、混合培养方式中培养物的稳定性、对既定环境中入侵物种的敏感性等。藻类在实验室中小规模培养与户外的大规模培养差异很大,因此,在测试藻种稳定性时,一定要进行户外大规模培养模拟实验。
2)建立藻种库
目前,还没有在世界范围内可以利用与维护的藻种信息库,首先私有企业的知识产权保护阻碍了藻种数据的统计,其次,关于藻种生长的详细信息要从藻类的培养过程中获得,这也是一项庞大到难以实现的工作。