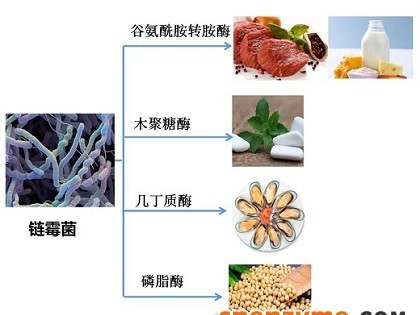
食品原料中内源酶对食品质量的影响及食品中加酶的作用
酶的作用对于食品质量的影响是非常重要的。实际上,没有酶或许就没有食品。对于任何一个生物体,酶参与了机体生长发育的每一个过程。食品原料的生长和成熟依赖于酶的作用,而在生物生长期间的环境条件影响着植物性食品原料的成分,其中也包括酶。
农产品的收获、贮藏和加工条件也影响食品原料中各类酶催化的反应,产生两类不同的结果,即可加快食品变质的速度,又可提高食品的质量。除了存在于食品原料的内源酶外,因微生物污染而引入的酶也参与催化食品原料中的反应。因此,控制酶的活力对于提高食品质量是至关重要的。本节将讨论影响食品颜色、质地、风味和营养质量的酶。www.cnenzyme.com
7.5.1 对颜色的影响
食品被消费者接受程度的如何,首先取决于食品的颜色,这是因为食品的内在质量在一般情况下很难判断。众所周知,新鲜瘦肉的颜色必须是红色的,而不是褐色或紫色的。这种红色是由于其中的氧合肌红蛋白所致。当氧合肌红蛋白转变成肌红蛋白时瘦肉就呈紫色。当氧合肌红蛋白和肌红蛋白中的Fe2+被氧化成Fe3+时,生成高铁肌红蛋白时,瘦肉呈褐色。在肉中酶催化的反应与其他反应竞争氧,这些反应的化合物能改变肉组织的氧化——还原状态和水分含量,因而影响肉的颜色。
绿色是许多新鲜蔬菜和水果的质量指标。有些水果当成熟时绿色减少而代之以红色、桔色、黄色和黑色。随着成熟度的提高,青刀豆和其他一些蔬菜中的叶绿素的含量下降。上述食品材料颜色的变化都与酶的作用有关。导致水果和蔬菜中色素变化的3个关键性的酶是脂肪氧合酶、叶绿素酶和多酚氧化酶。
7.5.1.1 脂肪氧合酶
脂肪氧合酶对于食品有6个方面的功能,它们中有的是有益的,有的是有害的。两个有益的是:小麦粉和大豆粉中的漂白;在制作面团中形成二硫键。四个有害的是:破坏叶绿素和胡萝卜素;产生氧化性的不良风味,它们具有特殊的青草味;使食品中的维生素和蛋白质类化合物遭受氧化性破坏;使食品中的必需脂肪酸,如亚油酸、亚麻酸和花生四烯酸遭受氧化性破坏。这6个方面的功能都与脂肪氧合酶作用于不饱和脂肪酸时产生的自由基有关。
7.5.1.2 叶绿素酶
叶绿素酶存在于植物和含叶绿素的微生物。它水解叶绿素产生植醇和脱植醇基叶绿素;尽管将果蔬失去绿色归之于这个反应,然而,由于脱植醇基叶绿素呈绿色,因此没有证据支持该观点。相反,有证据显示脱植醇基叶绿素在保持绿色的稳定性上优于叶绿素。
7.5.1.3 多酚氧化酶
多酚氧化酶又称为酪氨酸酶、多酚酶、酚酶、儿茶酚氧化酶、甲酚酶和儿茶酚酶。它主要存在于植物、动物和一些微生物(主要是霉菌)中,它催化食品的褐变反应。
7.5.2 对风味的影响
对食品的风味作出贡献的化合物不知其数,风味成分的分析也是有难度的。正确地鉴定哪些酶在食品风味物质的生物合成和不良风味物质的形成中起重要作用,同样是非常困难的。
在食品保藏期间由于酶的作用会导致不良风味的形成。例如,有些食品材料,像青刀豆、豌豆、玉米、冬季花椰菜和花椰菜因热烫处理的条件不适当,在随后的保藏期间会形成显著的不良风味。
在讨论脂肪氧合酶对食品颜色的影响时也提到它能产生氧化性的不良风味。脂肪氧合酶的作用是青刀豆和玉米产生不良风味的主要原因,而胱氨酸裂解酶(cystinelyase)的作用是冬季花椰菜和花椰菜产生不良风味的主要原因。下面介绍几种影响食品风味的酶。www.cnenzyme.com
7.5.2.1 硫代葡萄糖苷酶(glucosinolase)
在芥菜和辣根中存在着芥子苷(glucosinolates)。在这类硫代葡萄糖苷中,葡萄糖基与糖苷配基之间有一个硫原子,其中R为烯丙基、3-丁烯基、4-戊烯基、苯基或其他的有机基团,烯丙基芥子苷(allylglucosinolate)最为重要。硫代葡萄糖苷在天然存在的硫代葡萄糖苷酶作用下,导致糖苷配基的裂解和分子重排。生成的产物中异硫氰酸酯是含硫的挥发性化合物,它与葱的风味有关。人们熟悉的芥子油即为异硫氰酸烯内酯,它是由烯丙基芥子苷经硫代葡萄糖苷酶的作用而产生的。
7.5.2. 2 过氧化物酶
过氧化物酶普遍地存在于植物和动物组织中。在植物的过氧化物酶中,对辣根的过氧化物酶(horseradish peroxidase doner :hydrogen peroxideoxidoreductase EC 1.11.1.7)研究得最为彻底。如果不采取适当的措施使食品原料(例如蔬菜)中的过氧化物酶失活,那么在随后的加工和保藏过程中,过氧化物酶的活力会损害食品的质量。未经热烫的冷冻蔬菜所具有的不良风味被认为是与酶的活力有关,这些酶包括过氧化物酶、脂肪氧合酶、过氧化氢酶、α-氧化酶(α-oxidase)和十六烷酸一辅酶A脱氢酶。然而,从线性回归分析未能发现上述酶中任何两种酶活力之间的关系或任何一种酶活力与抗环血酸浓度之间的关系。
各种不同来源的过氧化物酶通常含有一个血色素(铁卟啉Ⅸ)作为辅基。过氧化物酶催化下列反应
ROOH + AH2 → H2O + ROH + A
反应物中的过氧化物(ROOH)可以是过氧化氢或一种有机过氧化物,例如过氧化甲基(CH3OOH)或过氧化乙基(CH3CH2OOH)。在反应中过氧化物被还原,而一种电子给予体(AH2)被氧化。电子给予体可以是抗坏血酸、酚、胺或其他有机化合物。在过氧化物酶催化下,电子给予体被氧化成有色化合物,根据反应的这个特点可以设计分光光度法测定过氧化物酶的活力。
目前对过氧化物酶导致食品不良风味形成的机制还不十分清楚,Whitaker认为应采用导致食品不良风味形成的主要酶作为判断食品热处理是否充分的指标。例如,脂肪氧合酶被认为是导致青刀豆和玉米不良风味形成的主要酶,而胱氨酸裂解酶是导致冬花椰菜和花椰菜不良风味形成的主要酶。然而,由于过氧化物酶普遍存在于植物中,并且可以采用简便的方法较准确地测定它的活力,尤其是热处理后果蔬中残存的过氧化物酶的活力,因此它仍然广泛地被采用为果蔬热处理是否充分的指标。
过氧化物酶在生物原料中的作用可能还包括下列几方面:①作为过氧化氢的去除剂;②参与木质素的生物合成;③参与乙烯的生物合成;④作为成熟的促进剂。虽然上述酶的作用如何影响食品质量还不十分清楚,但是过氧化物酶活力的变化与一些果蔬的成熟和衰老有关已经得到证实。
从前面的讨论中可以看出,食品原料中的一些内源酶的作用除了影响食品的风味外,同时还影响食品的其他质量,例如脂肪氧合酶的作用就同时影响食品的颜色、风味、质构和营养质量。在一些情况下几种酶的协同作用对食品的风味会产生显著的影响。
7.5.3 对质地的影响
质地是决定食品质量的一个非常重要的指标。水果和蔬菜的质地主要取决于所含有的一些复杂的碳水化合物:果胶物质、纤维素、半纤维素、淀粉和木质素。自然界存在着能作用于这些碳水化合物的酶,酶的作用显然会影响果蔬的质地。对于动物组织和高蛋白质植物性食品,蛋白酶作用会导致质地的软化。
7.5.3.1 果胶酶(见7.5.5.1水解酶类⑧)
7.5.3.2 纤维素酶
水果和蔬菜中含有少量纤维素,它们的存在影响着细胞的结构。纤维素酶是否在植物性食品原料(例如青刀豆)软化过程中起着重要作用仍然有着争议。在微生物纤维素酶方面已做了很多的研究工作,这显然是由于它在转化不溶性纤维素成葡萄糖方面潜在的重要性。
7.5.3.3 戊聚糖酶
半纤维素是木糖、阿拉伯糖或木糖和阿拉伯糖(还含有少量其他的戊糖和己糖)的聚合物,它存在于高等植物中。戊聚糖酶存在于微生物和一些高等植物中,它水解木聚糖、阿拉伯聚糖和阿拉伯木聚糖,产生相对分子质量较低的化合物。
小麦中存在着浓度很低的戊聚糖酶,然而对它的性质了解甚少。目前在微生物戊聚糖酶方面做了较多的研究工作,已能提供商品微生物戊聚糖酶制剂。
7.5.3.4 淀粉酶
水解淀粉的淀粉酶存在于动物、高等植物和微生物中。因此,在一些食品原料的成熟、保藏和加工过程中淀粉被降解就不足为奇了。由于淀粉是决定食品的粘度和质构的一个主要成分,因此,在食品保藏和加工期间它的水解是一个重要的变化。淀粉酶包括3个主要类型:α-淀粉酶、β-淀粉酶和葡萄糖淀粉酶。
α-淀粉酶存在于所有的生物,它从淀粉(直链和支链淀粉)、糖原和环糊精分子的内部水解α-1,4-糖苷键,水解产物中异头碳的构型保持不变。由于α-淀粉酶是内切酶,因此它的作用能显著地影响含淀粉食品的粘度,这些食品包括布丁和奶油酱等。唾液和胰α-淀粉酶对于消化食品中的淀粉是非常重要的。一些微生物含有高浓度的α-淀粉酶。一些微生物α-淀粉酶在高温下才会失活,它们对于以淀粉为基料的食品的稳定性会产生不良的影响。
β-淀粉酶存在于高等植物,它从淀粉分子的非还原性末端水解α-1,4-糖苷键,产生β-麦芽糖。由于β-淀粉酶是端解酶,因此仅当淀粉中许多糖苷键被水解时,淀粉糊的粘度才会发生显著的改变。β-淀粉酶作用于支链淀粉时不能越过所遭遇的第一个α-1,6-糖苷键,而作用于直链淀粉时能将它完全水解。如果直链淀粉分子含偶数葡萄糖基,产物中都是麦芽糖;如果淀粉分子含奇数葡萄糖基,产物中除麦芽糖外,还含有葡萄糖。因此β-淀粉酶单独作用于支链淀粉时,它被水解的程度是有限的。聚合度10左右的麦芽糖浆在食品工业中是一种很重要的配料。人体中的淀粉酶是一种巯基酶,它能被许多巯基试剂抑制。在麦芽中,β-淀粉酶常通过二硫键以共价方式连接至其他巯基上;因此,用一种巯基化合物(例如半胱氨酸)处理麦芽能提高它所含的β-淀粉酶的活力。
7.5.3.5 蛋白酶
对于动物性食品原料,决定其质构的生物大分子主要是蛋白质。蛋白质在天然存在的蛋白酶作用下所产生的结构上的改变会导致这些食品原料质构上的变化;如果这些变化是适度的,食品会具有理想的质构。
(1)组织蛋白酶(cathepsins) 组织蛋白酶存在于动物组织的细胞内,在酸性pH具有活性。这类酶位于细胞的溶菌体内,它们区别于由细胞分泌出来的蛋白酶(胰蛋白酶和胰凝乳蛋白酶),已经发现五种组织蛋白酶,它们分别用字母A、B、C、D和E表示。此外,还分离出一种组织羧肽酶(Catheptic carboxypeptidase)。
组织蛋白酶参与了肉成熟期间的变化。当动物组织的pH在宰后下降时,这些酶从肌肉细胞的溶菌体粒子中释放出来。据推测,这些蛋白酶透过组织,导致肌肉细胞中的肌原纤维以及胞外结缔组织例如胶原分解;它们在pH 2.5~4.5范围内具有最高的活力。
(2)钙活化中性蛋白酶(calsium-activated neutral proteinases CANPs) 钙化中性蛋白酶或许是已被鉴定的最重要的蛋白酶。已经证实存在着两种钙活化中性蛋白酶,即CANPⅠ和CANPⅡ,它们都是二聚体。两种酶含有相同的较小的亚基,相对分子质量约为30 000,及都含有不同的较大的亚基,相对分子质量约为80 000,在免疫特性方面有所不同,它们在结构上相符的程度约50%。尽管钙离子对于酶的作用是必需的,然而酶的活性部位中含有半胱氨酸残基的巯基,因此它归属于半胱氨酸(巯基)蛋白酶。
50~100 pmol/L Ca2+可使纯的CANPI完全激活,而CANPⅡ的激活需要1~2 mmol/L Ca2+,在CANPI被完全激活的条件下,CANPⅡ实际上是处在失活的状态。肌肉CANP以低浓度存在,它在pH低至约6时还具有作用。肌肉CANP可能通过分裂特定的肌原纤维蛋白质而影响肉的嫩化。这些酶很有可能是在宰后的肌肉组织中被激活,它们可能在肌肉改变成肉的过程中同溶菌体蛋白酶协同作用。
与其他组织相比,肌肉组织中蛋白酶的活力是很低的,兔的心脏、肺、肝和胃组织蛋白酶活力分别是腰肌的13、60、64和76倍。正是由于肌肉组织中的低蛋白酶活力才会导致成熟期间死后僵直体肌肉以缓慢地有节制和有控制的方式松弛,这样产生的肉具有良好的质构。如果在成熟期间肌肉中存在激烈的蛋白酶作用,那么不可能产生理想的肉的质构。
(3)乳蛋白酶 牛乳中主要的蛋白酶是一种碱性丝氨酸蛋白酶,它的专一性类似于胰蛋白酶。此酶水解β-酪蛋白产生疏水性更强的γ-酪蛋白,也能水解αs-酪蛋白.但不能水解κ-酷蛋白。在奶酪成熟过程中乳蛋白酶参与蛋白质的水解作用。由于乳蛋白酶对热较稳定,因此,它的作用对于经超高温处理的乳的凝胶作用也有贡献。乳蛋白酶将β-酪蛋白转变成γ-酪蛋白这一过程对于各种食品中乳蛋白质的物理性质有着重要的影响。
在牛乳中还存在着一种最适pH在4左右的酸性蛋白酶;然而,此酶较易热失活。
7.5.4 对营养价值的影响
有关酶对食品营养质量的影响的研究结果的报道相对来说较少见。前面已提及的脂肪氧合酶氧化不饱和脂肪酸确实会导致食品中亚油酸、亚麻酸和花生四烯酸这些必需脂肪酸含量的下降。脂肪氧合酶催化多不饱和脂肪酸氧化过程中产生的自由基能降低类胡萝卜素(维生素A的前体)、生育酚(维生素E)、维生素C和叶酸在食品中的含量。自由基也会破坏蛋白质中半胱氨酸、酪氨酸、色氨酸和组氨酸残基。在一些蔬菜中抗坏血酸氧化酶会导致抗坏血酸的破坏。硫胺素酶会破坏硫胺素,后者是氨基酸代谢中必需的辅助因子。存在于一些维生素中的核黄素水解酶能降解核黄素。多酚氧化酶引起褐变的同时也降低了蛋白质中有效的赖氨酸量。