
(广州博仕奥集团 510663)
摘 要:淀粉是谷物类饲料碳水化合物的主要成分,也是畜禽所需能量的重要能源。淀粉由于来源、组成及结构不同,其在畜禽消化道中消化的部位和速度存在明显差异,因而它们的供能效率也不同。淀粉在淀粉酶的作用下水解,不同类型的淀粉酶,其水解淀粉的效率、形成的产物各不相同。
关键词:淀粉 直链淀粉 支链淀粉 淀粉酶 脱支酶
1 淀粉结构
淀粉(Starch)是由许多葡萄糖分子缩合而成的多糖,化学成分是:C44.4%,H6.2%,O49.4%,化学式(C6H10O5)n。
作为能量储藏形式的淀粉,广泛存在于高等植物当中,按结构和来源可分为A型的谷物淀粉(来自玉米、小麦、水稻、大麦、糯米等);B型的块根块茎淀粉(来自木薯、甘薯、马铃薯);C型的豆类种子淀粉(来自大豆、豌豆、蚕豆),其他淀粉(如西米)(姚新灵,2001)。
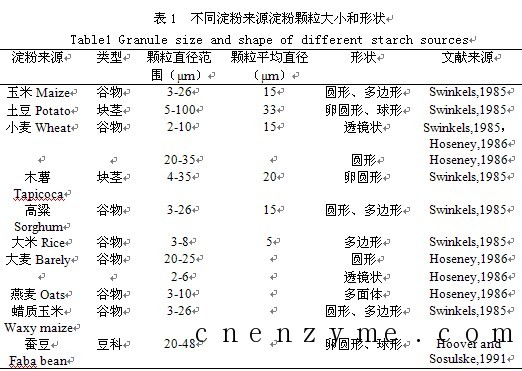
淀粉在植物体内以淀粉粒的形式存在,胚乳造粉体是淀粉粒形成的地方。不同植物来源,淀粉颗粒形态和大小各异,淀粉颗粒的直径从不足1μm到100μm,形状有球形、卵圆形、多角形、混合形等多种,见表1。
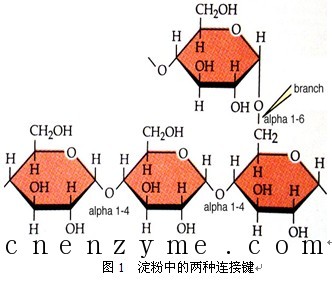
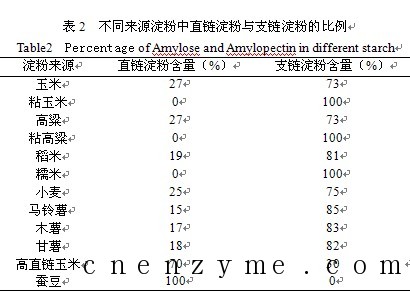
淀粉颗粒虽然只含有葡萄糖一种单糖,但其成分并不是均一的,它可以用热水分成两部分:一部分可溶解于热水,水溶液粘度小,称为直链淀粉(Amylose);另一部分难溶于热水,水溶液成糊状,粘度较高,称为支链淀粉(Amylopectin)。淀粉中的葡萄糖之间的连接存在两种键,一种是α-1,4糖苷键,主要见于直链淀粉;另外一种是α-1,6糖苷键,主要见于支链淀粉,见图1。天然的淀粉主要由直链淀粉和支链淀粉组成,淀粉的来源不同,两种组分的比例也不同。大多数淀粉含直链淀粉20%~25%,含支链淀粉75%~80%,见表2。
1.1 直链淀粉
直连淀粉是由D-葡萄糖通过α-1,4糖苷键聚合而成的直线型大分子,由1000-5000个葡萄糖单元连结而成,基本不分支或很少分支,分子量为105-106kDa。直链淀粉主要通过分子内氢键,使长链分子卷曲形成螺旋形的构象而存在,螺旋的每一圈含有6个葡萄糖单元。这种螺旋构象,在分子链上各极性基团的相互作用下产生弯曲与折叠。
1.2 支链淀粉
支链淀粉是多数淀粉的主要成分,支链淀粉除含有α-1,4糖苷键外,还存在许多α-1,6糖苷键的分支,如树枝状,分子量比直链淀粉要大,约为107-109kDa,由5000-50000个葡萄糖单元构成。支链淀粉分支较多,平均20-26个α-1,4糖苷键连接的葡萄糖残基就有一个α-1,6糖苷键分支点。支链淀粉共有A 、B 、C 三种糖链,C 链为支链淀粉的骨架主链,具有一个还原性末端,每个分子只有一个C 链;B 链是经α-1,6- 糖苷键与C 链相连接的分支链;A 链是通过还原性末端与其他链连接的链,是淀粉分子最外侧不含分支的链。A 链与B 链的比值实际上反映了支链淀粉分子的分支化度,分支化度定义为每条B链上所具有的A 链数目(Marshall JJ,1974),见图2。因此,支链淀粉与直链淀粉在物理、化学性质上存在一些差异,见表3。

1.3 直/支链淀粉比对畜禽生产性能的影响
淀粉颗粒大小、非淀粉多糖、日粮油脂水平、内源淀粉消化酶活性、直/支链淀粉比等,都影响畜禽对淀粉的消化率(Cummings J H,1995)。
宾石玉等(2006)研究不同日粮来源的淀粉(玉米、早籼稻糙米和糯米)对断奶仔猪生产性能的影响,发现直链淀粉和支链淀粉的比值大小显著影响断奶仔猪的生产性能,其中玉米组(直/支比为0.23)日增重比糯米组(直/支比为0)高24.53%,料肉比低18.18%。
戴球仲等(2008)分别用直/支链淀粉比为0.11、0.23、0.35、0.47的日粮饲喂黄羽肉仔鸡,结果表明,直/支链淀粉比为0.23的日粮组料肉比最低,日粮大多数氨基酸消化率最高,日粮直/支链淀粉比可能改变消化道中葡萄糖的供给状态,从而影响饲料蛋白质和氨基酸的消化道代谢。
2 饲用外源淀粉酶系
淀粉酶(Amylase)的主要功能是水解淀粉,按其水解淀粉的作用方式不同分为α-淀粉酶(α-Amylase)、β-淀粉酶(β-Amylase)、糖化酶(葡萄糖淀粉酶Glucoamylase)、脱支酶(Debranchins enzyme)。
2.1 α-淀粉酶
α-淀粉酶,也称为内切淀粉酶,主要是水解淀粉分子内部的α-1,4糖苷键,且水解此糖苷键的次序是无规律的;而不能水解淀粉分子内部的α-1,6糖苷键,但可跨越此键水解分子内部的α-1,4糖苷键。α-淀粉酶水解直链淀粉的最终产物是麦芽糖和葡萄糖,而水解支链淀粉的最终产物是麦芽糖、葡萄糖、异麦芽糖和含有α-1,6糖苷键的极限糊精。
2.2 β-淀粉酶
β-淀粉酶,也称为外切淀粉酶,该酶从淀粉分子的非还原性末端开始,水解相间隔的α-1,4糖苷键,依次切下麦芽糖单元,同时发生瓦尔登转位反应(Walden inversion),使产物由α-型转变为β-型麦芽糖。β-淀粉酶只能水解α-1,4糖苷键,而不能水解α-1,6糖苷键,且不能跨越此键,遇到此键水解作用停止。β-淀粉酶水解直链淀粉时,麦芽糖的生成速度较慢;而水解支链淀粉时,因支链淀粉分枝较多,非还原性末端较多,所以麦芽糖生成速度较快。但β-淀粉酶不能水解和跨越α-1,6糖苷键,所以只能水解分支点以外的部分,产生的麦芽糖相当于支链淀粉总量的50-60%,而不能水解分支点以内的部分,此剩余部分一般称为极限糊精(孟庆红,1997)。
2.3 糖化酶
糖化酶从淀粉分子的非还原性末端开始,逐次切下葡萄糖。糖化酶不仅能水解α-1,4糖苷键,而且能水解α-1,6糖苷键和α-1,3糖苷键,但水解后两者的速度要较前者慢很多。所以,糖化酶水解直链淀粉和支链淀粉时,能将它们全部水解为葡萄糖,只是时间和效率的问题。
2.4 脱支酶
脱支酶是一种能够高效、专一性地切开支链淀粉分支点α-1,6-糖苷键,从而剪下整个侧枝,形成直链淀粉的一种酶。由于淀粉中支链淀粉含量相对较高,但其他淀粉酶都不能水解支链淀粉中的α-1,6-糖苷键或者水解效率很低,所以淀粉中的α-1,6-糖苷键成为淀粉水解中的一个阻碍性因素。α-淀粉酶、β-淀粉酶以及二者共同作用均不能将淀粉彻底水解为小分子糖,而需要脱支酶的参与。所以,添加脱支酶,能有效提高淀粉的利用率。
根据底物特异性的差异,脱支酶分为几种(周瑞芳等,1993):低聚葡萄糖苷酶(Oligo-1,6-glucosidase)、淀粉-1,6-葡萄糖苷酶(Amylo-1,6-glucosidase)、异淀粉酶(Isoamylase)、普鲁兰酶(Pullulanase)。脱支酶的应用主要以普鲁兰酶和异淀粉酶较多,但普鲁兰酶和异淀粉酶对底物的特异性存在差异。当分支点处葡萄糖残基数量大于二时,普鲁兰酶就可切断α-1,6 - 糖苷键,其作用底物的最小单位为麦芽糖基麦芽糖。而异淀粉酶在分支点的葡萄糖残基少于三个时就不能发挥作用,其作用底物的最小单位为麦芽三糖基麦芽四糖(Marshall JJ,1974)。

2.4.1 低聚葡萄糖苷酶
该酶不能直接分解淀粉中的α-1,6-糖苷键,而只能分解经α-淀粉酶水解后的α—糊精中的α-1,6-糖苷键。
该酶是一种典型的间接脱支酶,可作用于外露一个葡萄糖残基的α-1,6-糖苷键。
这是切断支链淀粉及某些分支糊精中的α-1,6-糖苷键的酶,对普鲁兰糖则完全没有作用。
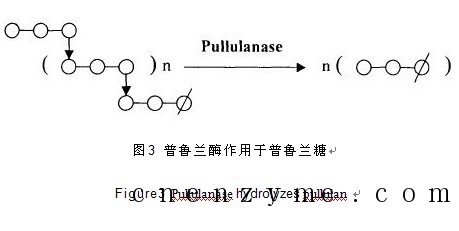
1961 年Bender H.等人在研究一株产气杆菌Aerobacter aerogenes ( 现名klebsiellapneumon iac)时,首次发现普鲁兰酶。该酶能专一性水解α-1,6-糖苷键,使支链淀粉型多糖的分支链脱离主链,主要作用的底物有:普鲁兰糖(Pullulan)、支链淀粉和β-极限糊精,最终产物是麦芽三糖。普鲁兰糖亦称茁霉多糖,其结构是葡萄糖按α-1,4-糖苷键结合成麦芽三糖,两端再以α-1,6-糖苷键同另外的麦芽三糖结合,并以这种连接方式形成的高分子多糖。普鲁兰酶在改善淀粉酶对淀粉的作用效果,提高淀粉利用率中有着重要作用,普鲁兰酶协同α-淀粉酶、糖化酶,能将淀粉彻底降解为葡萄糖。在脱支酶中,普鲁兰酶要求的底物分子结构最小,能将最小单位的支链分解,所以应用最为广泛。

3 畜禽对淀粉的消化利用
在畜禽机体中,主要的淀粉酶有唾液淀粉酶(Salivary amylase)和胰淀粉酶,均属于α-淀粉酶。食物在畜禽口腔停留的时间一般不长,所以唾液淀粉酶对淀粉的消化非常有限,淀粉在胃内的消化也是比较少。淀粉在畜禽消化道水解的主要部位是小肠,小肠中的α-淀粉酶把淀粉中的直链淀粉分解为麦芽糖和麦芽三糖,支链淀粉分解为麦芽糖、麦芽三糖和极限糊精。胰腺和肠粘膜产生的麦芽糖酶、蔗糖酶可以继续分解这些糖类。由于畜禽体内缺乏水解α-1,6糖苷键的酶,所以支链淀粉中的α-1,6糖苷键成为淀粉充分消化吸收的障碍,淀粉在畜禽机体消化并不完全。
4 脱支酶能有效提高淀粉利用率
目前在饲料酶中,应用较广的外源淀粉酶主要有α-淀粉酶和糖化酶,但对脱支酶的研究与应用报道较少。刘小英(2009)研究支链淀粉酶对生长蛋鸡生产性能的影响,发现在饲料中分别添加
在淀粉糖工业生产上,利用脱支酶,配合α-淀粉酶和糖化酶的使用,能显著提高淀粉的水解率。由于畜禽机体内部缺乏水解支链淀粉α-1,6糖苷键的酶,而玉米、小麦、木薯等作为主要能量饲料,在畜禽日粮中比例较大、支链淀粉的含量相对较高,所以脱支酶的使用,必将大大提高淀粉的消化率。
(参考文献略。需者可函索)




