
到2020年,饲料用粮预计达到2.98亿t,将取代口粮在中国粮食用途中占最大份额[1]。人畜争粮现状是我国正面临的严峻挑战之一,因此,开发新型饲料和提高饲料的利用水平势在必行。多酶生物饲料是一种通过生物技术的手段使饲料中富含多种酶类的饲料,与普通饲料相比,其所含的酶类能够在一定程度上消除饲料原料中抗营养成分或者对内源性酶类起到补充作用,从而在一定程度上提高饲料的利用水平。
1、饲用酶及应用效果的研究
饲料酶制剂大致可分为非消化酶和外源性消化酶两大类。非消化酶是指动物自身无法分泌到消化道内的酶,这类酶能消化动物自身不能消化的物质或降解一些抗营养因子,主要包括纤维素酶、木聚糖酶、β-葡聚糖酶、植酸酶、果胶酶等等。外源消化酶是指通过技术手段外加的消化酶,这些酶动物自身可以分泌,如淀粉酶、蛋白酶和脂肪酶类等。对主要饲用酶做如下介绍。
1.1木聚糖酶
木聚糖酶主要是指β-1,4-D-木聚糖酶,它以内切的方式水解木聚糖分子中的β-1,4-木糖苷键,其水解产物有低聚糖和少量木糖等单糖。木聚糖酶在饲料中的主要作用在于降解木聚糖,从而降低食糜黏度。此外,低聚木糖还能调节动物肠道微生态环境、减少动物结肠炎的发生以及降低抗生素等兽药用量的功效[2]。Tapingkae研究了酸性木聚糖酶对小猪的消化吸收以及产仔情况的影响,发现饲喂6周后的断奶小猪日增重相对提高63g/d,腹泻率降低50%[3]。Vandeplas等在肉鸡日粮中添加不同来源的木聚糖酶,肉鸡日增重相对提高4.5%,粗纤维消化吸收率提高58.9%
[4]。此外,Vandeplas的研究还表明,饲料中加入木聚糖酶后,肠道中双歧杆菌的数量明显增加,同时粪便中鼠伤寒沙门氏菌的数量明显降低[5]。
1.2β-葡聚糖酶
β-葡聚糖酶是一类能催化β-葡聚糖水解的多种酶类的总称。其中β-1,3-1,4-葡聚糖酶活性为最高。β-葡聚糖酶能够水解饲料原料中的β-葡聚糖,生成寡糖和葡萄糖;消除由于β-葡聚糖的高吸水、高持水等特性而产生的物理作用,从而降低消化道内容物黏度,提高禽畜消化吸收水平。Newman等人在大麦为基础的饲料中添加β-葡聚糖酶,仔猪日增重和饲料转化率分别提高1%和16%[6]。Campbell等在猪饲料中添加β-葡聚糖酶,能量和蛋白质利用率分别提高13%和21%[7]。
1.3β-甘露聚糖酶
β-甘露聚糖酶是一类能够催化含有β-1,4-甘露糖苷键的甘露聚糖及其衍生物水解的内切水解酶[8]。甘露聚糖酶在饲料中的作用在于能够降解饲料原料中的甘露聚糖,降低食糜的黏度,解除甘露聚糖与金属离子、消化酶、胆盐的结合作用,增加养分的吸收;并且有研究表明,酶解生成的甘露低聚糖可用作双歧杆菌促生长因子[9],使有益菌繁殖,对动物肠道起着重要的调节作用。另外,甘露寡糖还能够刺激免疫应答,增强动物细胞免疫和体液免疫[10]。
张辉华等研究β-甘露聚糖酶对肉鸭椰籽粕消化率的影响,结果表明添加β-甘露聚糖酶后蛋白质消化率与真实消化率分别提高8.04%与7.35%;能量利用率与真实利用率分别提高3.61%与3.38%[11]。乔海云等研究表明,在玉米-豆粕型日粮中添加β-甘露聚糖酶可以提高肉鸡的免疫力,降低肠道pH
值,抑制大肠杆菌繁殖,增加有益菌的数量[12]。
1.4α-半乳糖苷酶
α-半乳糖苷酶是水解α-半乳糖苷键的一种外切糖苷酶,能专一性催化多糖、糖脂、糖蛋白中糖链末端的α-1,6半乳糖苷键水解,其不仅能水解棉籽糖和水苏糖等低聚糖,还能水解含α-半乳糖苷的杂多糖[13]。在饲料中添加α-半乳糖苷酶的作用在于能够降解饲料原料中的α-半乳糖苷,破坏食糜的水化膜使食糜蠕动减缓,提高食糜消化程度。同时还可抑制肠道内微生物的过度发酵,使胺和甲酚等有害物质和气体减少,提高内源性蛋白酶的活力,增加多肽类物质,控制因饲料消化不良引起的腹泻[14]。蒋小丰等研究表明,玉米-豆粕型日粮中添加α-半乳糖苷酶能提高断奶仔猪对蛋白质的利用率,促进抗体合成,提高粗纤维、粗脂肪和粗蛋白质的表观消化率[15]。Baucells等研究表明,在肥育猪日粮中添加α-半乳糖苷酶能分别提高干物质消化率和蛋白质消化率2.8%和12.5%[16]。戴求仲等研究表明,在玉米-豆粕型日粮中添加α-半乳糖苷酶能够提高仔猪的平均日增重,明显降低仔猪腹泻率[17]。
1.5果胶酶
果胶酶是指是能够分解果胶质的多种酶的总称,可以裂解β-1,4-糖苷键,有效地分解植物细胞壁间的果胶质,破坏细胞组织。果胶酶在饲料中的作用在于降解果胶,降低肠道食糜的黏度,还能使植物细胞壁、液泡等受损,提高饲料营养和能量的利用率。姜晓霞研究结果表明,饲料中添加果胶酶能显著提高肉鸡的平均日增重和体重[18]。Tahir等研究表明,复合使用果胶酶与纤维素酶显著提高肉鸡的屠宰率、胸肌率[19]。梅宁安等研究也表明日粮中添加果胶酶能显著提高肉仔鸡的生长性能[20]。
1.6植酸酶
植酸酶即肌醇六磷酸水解酶,是水解植酸及植酸盐成肌醇与磷酸的一类酶的总称。植酸酶在饲料中的作用,主要体现在可以促进植酸的消化和吸收,从而在一定程度上消除其抗营养作用。在动物体内,由于植酸是螯合剂,所以会引起禽畜出现微量元素缺乏症;植酸也是许多蛋白酶的抑制剂,从而影响禽畜的消化功能。另外,植酸还能够和蛋白质结合为植酸复合体,降低蛋白质的消化利用率。单胃动物体内缺乏植酸酶,无法利用植酸,80%以上的磷从动物粪便中排出,从而造成环境的污染和资源的浪费。Powell等在低磷日粮中添加植酸酶,可以提高肉鸡生长速度以及钙、磷消化率和钙利用率[21]。Pilial等研究发现,在肉鸡低磷日粮中添加植酸酶,可显著提高屠宰率[22]。
2.7纤维素酶
纤维素酶是一类能够降解纤维素为葡萄糖的多组分酶系的总称,它们协同作用,分解纤维素产生寡糖和纤维二糖,最终水解为葡萄糖。纤维素酶在饲料中的作用主要在于:对植物原料的细胞壁进行降解,促进营养的吸收;增强机体代谢水平和机体免疫力,有利于改善动物微生态环境。邓玉英等在断奶羔羊日粮中添加0.5%纤维素酶,结果羊平均日增重提高4.22%(P>0.05),料重比降低5.20%,并且腹泻率明显降低[23]。Eun等在以青贮玉米和苜蓿干草为主的饲料中添加纤维素酶和木聚糖酶饲喂奶牛,中性洗涤纤维的降解度提高[24]。毛倩倩等研究表明,高纤维饲粮中添加果胶酶能够提高五龙鹅的生长和屠宰性能,果胶酶与纤维素酶配合使用效果更好[25]。
2.8外源消化酶
饲料中淀粉、蛋白质等大分子化合物是禽畜的营养来源,动物体内可以分泌相应的内源性消化酶,比如淀粉酶、蛋白酶、脂肪酶等等,但是由于动物生理上的差异,不同的动物消化道中的酶系不同,特别是幼龄动物或动物处于病态和应激情况下,酶的分泌较弱,此时,在饲料中添加消化酶可以起到对内源性消化酶补充的作用,并且也可以促进一些较难水解的营养组分的降解,比如血粉蛋白、膨化羽毛粉蛋白等,进而提高饲料的利用率和畜禽的生产性能。
何前等在黄羽肉鸡饲料添加脂肪酶,能显著提高饲料中粗脂肪的表观利用率[26]。Angle等在低蛋白日粮中添加外源蛋白酶后,在肉鸡生产性能上,低蛋白日粮组与正常蛋白日粮组无显著差异,饲料转化效率相对提高[27]。
3、多酶饲料制备工艺的研究
多酶饲料制备的工艺主要有以下两种:加酶法,即通过外加现成的饲料酶的方式制备多酶饲料,这一种方式较为常见;发酵法,即利用微生物发酵产酶的方式制备多酶饲料。联合使用外加酶的酶解作用和微生物的发酵作用的方式制备多酶饲料,也归为发酵法一类。
3.1加酶法
加酶法制备多酶生物饲料的工艺简单,是目前较为常见的方法,对于酶制剂的选择,不是简单的复配,而是要根据饲料原料的不同以及酶种类的特性有机组合。比如,用于豆粕型饲料的饲用酶以α-半乳糖苷酶为主,同时需要添加木聚糖酶等酶类;而大麦型饲料则主要以β-葡聚糖酶为主。我国农业部对饲料酶的添加也做了相应的规定与建议,见表1。

3.1.1加酶法多酶生物饲料的制备工艺
提高酶制剂的稳定性是加酶法制备多酶生物饲料的主要研究内容。由于在饲料的加工过程中,会经历许多条件较为剧烈的阶段,比如,制粒、膨化等阶段,在这些阶段中,饲料会受到温度、压力和湿度等的强烈作用,会使酶活严重损失甚至完全丧失,严重影响多酶饲料的质量。有关研究表明,通过挤压膨化工艺后,酶活会完全丧失,通过环模制粒工艺后,酶活力的损失也非常明显[28]。另外,酶作为一种蛋白质,动物的消化作用对其活性的影响也很大,所以对所加饲用酶酶活的保护是一个棘手的技术问题。目前常用确保酶活力的制备工艺有两种:一是利用载体或包被的稳定性技术,主要采用物理包埋、化学修饰来提高酶的稳定性以在饲料加工过程减少酶活的损失。如将液态饲用酶吸附在特定载体上,或将颗粒状的酶制剂用包被剂包被起来,可使这些饲用酶制剂在经历高温阶段后保持较高的活性,并可减少动物肠道对酶活力的不良影响。二是制粒后喷涂,这种技术是待饲料制粒后,将酶以液态的形式通过一定设备喷涂于其表面,从而避免了在饲料加工后期工序中酶活力的损失。
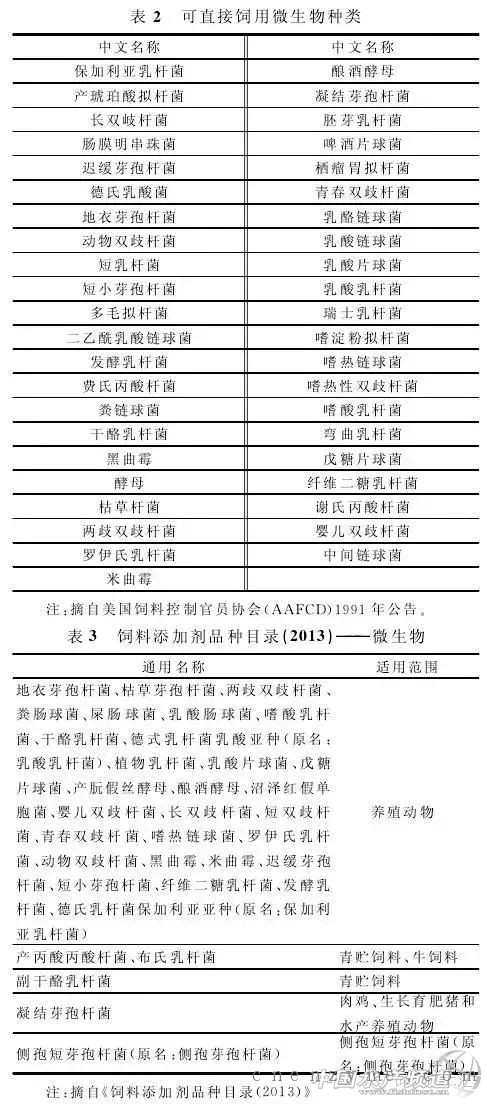
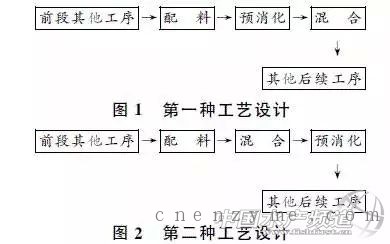
加酶法的另一种方式是体外消化方式,章世元等提出两种工艺流程设想:一是配料结束,混合前加入预消化工序(图1);二是混合结束后,制粒、膨化等后续其他工序前加入预消化工序(图2)[29]。此外,章世元等还研制了两种体外消化设备:即釜式调质器和改造型混合机。经过试验证明,体外预消化生产工艺可促进饲用酶发挥作用,两种预消化生产工艺可显著改善加酶饲料质量及营养品质,无机磷含量和还原糖含量显著提高,并且酸性洗涤纤维明显降低[30]。
章世元等通过实验得出加酶饲料在含水量14.5%、温度45℃条件下作用30min预消化效果较好,酸性洗涤纤维含量相对降低9.64%[31]。徐炜研究表明,加酶饲料经预消化处理后明显提高了动物对饲料养分的消化代谢率。与常规加酶饲料比,预消化加酶饲料的鸡饲料能量表观代谢率、蛋白质表观代谢率和磷表观代谢率分别提高6.20%、12.45%和67.39%[32]。
3.1.2加酶法制备多酶生物饲料存在的问题
虽然在加酶法制备多酶饲料的研究中,研究人员采取各种方法解决酶活问题,但是在加酶法制备多酶饲料工艺上,仍有很多不足之处,需要进一步研究和改进。载体法、包被法和喷涂法虽然在一定程度上防止了饲料加工过程中和动物消化道中不利因素的影响,但是并不能绝对确保酶制剂在后处理过程中的稳定性。比如,被包被的饲用酶,经过90℃通蒸汽加热会使酶迅速失活[33]。当制粒的调质温度为65℃时,载体对酶制剂有一定的保护作用,而当温度为75℃时,将失活70%左右[34]。另外,由于目前饲用酶基本为微生物所产,酶解的最适条件与动物体内条件不一定相符,而且动物体内环境复杂、多变,不同动物、不同生长发育阶段的动物体内的环境也不一样,从而饲用酶的体外测量和体内实际作用存在出入。最后,要将技术推广使用,成本也是一个非常重要的因素,无论是喷涂法、包被法还是载体法都会极大增加饲用酶的生产和应用成本。
体外消化加酶法虽然没有上述存在的问题,但是其消化工艺参数的有效控制比较困难,而且消化步骤所投入的时间较长,设备成本也较高。
3.2发酵法
发酵法制备的多酶生物饲料,属于发酵生物饲料的一种。利用微生物产酶,不仅可以得到多种饲用酶,起到改善饲料品质,提高利用率的效果,还能通过调节动物体内的微生态平衡,达到促进免疫力、增强机体机能等功效。
3.2.1菌种
利用微生物发酵法制备多酶生物饲料,首要关键点在于菌种的选择,好的菌种,生存能力强,产酶活力高,在饲料中能够起到更好的效果。除此之外,在饲料中添加的微生物,还需要满足众多其他条件,比如所选用的微生物,最好是来源于家畜体内,这样可以在一定程度上确保微生物进入动物体内后,不会因为微生物本身或者所产的其他物质致使动物患病或者死亡,确保安全性。另外这些微生物也更容易在家畜体内定植和繁殖并进行新陈代谢,从而发挥作用。而使用外源性微生物就必须要通过实验动物所做的急性、亚急性毒性实验和致畸致残等试验。另外,如果采取多菌种混合发酵,则要求各菌种在生理条件和产酶条件上能具有协同性,并且所产酶类能具有互补性。如果所用菌种是益生菌株,还要考虑到菌体的抗逆性,尤其需要对胆酸盐和胃酸具有较强的抵抗能力,这样才能够在进入禽畜的消化道后继续发挥作用,起到益生的效果。
美国FAD 批准用作直接饲喂的微生物已有43种(表2)。目前,我国农业部也在《饲料添加剂品种目录(2013)》(表3)中公布了可在饲料中添加的微生物种类,并于2014-02-01开始实施。
3.2.2发酵法多酶生物饲料的制备工艺
发酵法制备多酶饲料与加酶法相比,由于加入发酵步骤,在工艺上要复杂一些,但所含有的酶为微生物代谢产生,所以能够在一定程度上降低由于外加酶所带来的成本。针对单菌种或者多菌种混合固态发酵制备多酶饲料的工艺,主要有以下几个技术关键点:含水量需要控制得当。固体发酵与液态发酵的主要区别在于含水量,含水量会影响到菌体的生长代谢以及产物的形成。含水量多少应根据所选菌种及原料的不同来确定,高含水量会增加基质粘度,容易结块,影响透气和散热,也不利于基质中营养物质的利用与抗营养因子的水解;含水量低时,微生物生长受到抑制,尤其是真菌菌丝的生长和孢子的萌发,并且培养基表面很快干燥,发酵不彻底,致使酶活力降低。在发酵过程中,翻料时可以适量加入无菌水以及时补充水分。菌种的接种量应严格控制,接种量太大,则会出现基质消耗过快,使热量与废气堆积,产酶能力较低。对于霉菌而言,发酵后期还会出现孢子现象,不利于抗营养因子的水解;接种量太小,菌体量不够大,不利于产酶,并且发酵周期延长,污染杂菌的几率增加。相比单菌种发酵,多菌种混合发酵的条件控制较为困难,不同菌株的生长代谢会存在或大或小的差异,但是混合发酵可以有效地利用它们之间的互补性。对于多菌种混菌发酵而言,接种顺序与比例对多酶饲料的制备影响很大。
如王慧杰等人在制备含酶蛋白饲料的研究中将糖化和发酵分步进行。因为这样做可以使各菌种均可以在最佳条件下进行作用,而同步进行的糖化和发酵都很难在最佳条件下进行作用[35]。如果采用酶菌共效的方式,还需要考虑到酶的添加时间与用量,使得酶解作用能够与菌种的发酵作用相协调。温度也是需要考虑的重要因素,温度的变化不仅会影响微生物的生长、代谢以及培养基的传质、黏度等;还会影响产物酶的稳定性。发酵温度过高,不仅耗能较多,而且会加剧基质水分的蒸发,不利于产酶。固态物料的厚薄对发酵过程也有较大的影响。培养基太厚,不利于气体传递,会抑制微生物的生长,从而酶活性较低。另外,pH值也是一个很重要的参数,在固态发酵过程中较难控制,培养基中氮源对其影响较大,比如,铵盐做主要氮源时,易引起基质酸碱度下降。在培养基质中,还有很多主要组分以外的成分对酶的活力也有一定的影响。比如无机盐类、辅料等等。麦麸是一种很好的饲料原料,也是一种很好的发酵辅料,能够起到膨松和辅助氮源的作用,从而促进微生物生长和产酶,但麸皮过多也会使基质黏性增加,不利于透气,影响微生物的生长。对于无机盐类也需要注意,有时对酶活力影响很大,比如,Zn2+、Ca2+
和Cu2+ 对木聚糖酶有微弱的促进作用,Fe2+、Fe3+ 和Mn2+ 对木聚糖酶明显抑制[36]。
卫洋洋等用黑曲霉发酵制备油茶饼粕多酶生物饲料。确定发酵最佳基质中油茶饼粕与豆粕的混合比例为1︰1,料水比为1︰0.5,添加1%的硫酸铵,0.1%的磷酸氢二钾。结果表明,以pH5,温度28℃,1.5%接种量为最佳发酵条件,培养4d后,纤维素酶活力达到1 481.9
U/g,蛋白酶活力达到4 128.1U/g,蛋白质相应提高,各种抗营养因子如粗纤维。单宁等明显降低[37]。徐海燕,张志焱等以啤酒糟为主要原料,利用黑曲霉固态发酵生产富酶蛋白饲料。结果表明:最适发酵培养基啤酒糟∶棉粕比例为5∶5,KH2PO40.2%;最适发酵条件为:料水比为1∶1,接种量为含4.0×106
个孢子的孢子悬液,发酵温度为30℃,培养时间为54~60h,纤维素酶达到1 991U/g,木聚糖酶达到1 998U/g,酸性蛋白酶达到4 559U/g[38]。庄童琳等采用两株黑曲霉对苹果渣进行固态混菌发酵生产多酶生物饲料。结果表明:采用苹果渣︰棉粕比例1︰1,1%(NH4)2SO4,0.1%KH2PO4为最佳培养基,两株黑曲霉最佳配比为2︰1,接种量0.4%,30℃恒温培养48h,蛋白酶和纤维素酶活力分别为3 674、1 207U/g,果胶与单宁的降解率分别为99.0%和66.1%[39]。陈曦等采用米曲霉和酵母菌,利用混合酒糟制备含酶蛋白饲料。结果表明:最优的培养基配比是啤酒糟︰黄酒糟︰豆粕︰麸皮为4︰1︰1︰4,初始料水比为1︰1.8,硫酸铵2%和磷酸盐1%,米曲霉在酵母菌培养16h时接入,发酵周期为68h。混株发酵与单株发酵相比,酵母数提高了3倍,平均粗蛋白质提高5.32%,粗蛋白质增幅为35.74%,且发酵产物中含有蛋白酶901U/g,纤维素酶1 824U/g[40]。
3.2.3发酵法制备多酶生物饲料存在的问题
目前发酵法多酶饲料的研制与开发已取得了较大进步,但是仍存在许多技术问题。在菌种方面,由于添加到饲料中的微生物必须保证其安全性,所以大都采用没有经过改良的原始菌种作为发酵菌种,所以其酶活力一般比较低,这是发酵法的主要技术问题。另外,采用混合菌种时,如何很好地利用各菌种的互补性,这一方面也尚未研究透彻。在工艺方面,发酵法存在发酵速度慢,周期长,工作强度大,较难自动化等问题。由于发酵过程传质不均匀,天然原料成分复杂多变,菌体生长和酶的产生在各处也不均一,使得发酵参数的检测和控制也比较困难,从而影响到饲料的质量。
4、展望
多酶生物饲料的开发不仅能够提高现有饲料的利用水平,更能够扩大可利用饲料原料的范围,为新饲料原料的开发与利用提供新途径,对促进养殖业的发展有重要意义。就目前而言,对于多酶生物饲料的研发还存在许多问题,未来还需要在酶活力的保护与提高、酶类与原料的选择、禽畜消化道的适应性、混菌发酵的协调性、菌种的益生性等方面多做研究工作和加大投入,争取取得更有突破性的解决方案。多酶生物饲料是饲料开发领域不容置疑的发展方向,相信会有更多种类、更多功效的多酶生物饲料被开发成功。




